Great American carbonate bank, biostratigraphy
| The great American carbonate bank: The geology and economic resources of the Cambrian-Ordovician Sauk megasequence of Laurentia | |

| |
| Series | Memoir 98 |
|---|---|
| Chapter | Biostratigraphy and chronostratigraphy of the Cambrian-Ordovician great American carbonate bank |
| Author | J. F. Taylor, J. E. Repetski, J. D. Loch, S. A. Leslie |
| Link | Web page |
| PDF file (requires access) | |
| Store | AAPG Store |
In their synthesis of Cambrian biostratigraphy in North America, Lochman-Balk and Wilson[1] discussed in detail the nature of the faunal units that they used to subdivide and correlate strata that formed in shallow-marine environments of the continent (their cratonic and intermediate biofacies realms) in the early Paleozoic. They made clear the practical and conceptual differences between a teilzone through which a species or genus ranges locally and a biozone, an extensive time-rock unit that approximates the total span of time through which a particular genus survived. The greater geographic extent of the genus-based biozone rendered it more useful for interregional correlation. By expanding the concept to define a time-rock unit representing the total stratigraphic range of any of several characteristic genera or species in a stratigraphically significant assemblage, which they termed the faunizone, they had some success in correlating into coeval deep-water successions (their extracratonic euxinic realm). A similar assemblage-zone concept was used in the definition and correlation of the original alphanumeric zones established on the stratigraphic ranges of broadly defined genera and species of trilobites[2] [3] and conodonts[4] in the Lower Ordovician.
Numerous studies through the latter half of the 20th century yielded considerable refinement of the early coarse biostratigraphic framework, producing many species-based subzones within the genus-based assemblage zones. Numbered rectangles are provided in Figure 1 and Figure 2 to provide additional references that contain detailed information on the zones and subzones recognized in particular areas of North America. These typically include detailed range charts that show the stratigraphic distribution of species recovered from measured sections and from drill core. They also generally provide a complete listing of the species that occur within each zone or subzone and identify the species whose lowest occurrence(s) define(s) the base of the unit. It became fairly standard practice to leave the zones topless by simply defining the top of each unit as the base of the overlying unit. By this convention, barren intervals are assigned to the top of the subjacent zone or subzone. This practice has the disadvantage in studies of subsurface material recovered from well cuttings of emphasizing lowest occurrences (FADs) though highest documented occurrences (last appearance datums) are less prone to distortion by downhole transport of material by caving or fluid circulation. This distortion is less of a problem in data collected from core material, as is typically the case for macrofossils.
In concept, the boundaries of biozones and subbiozones are independent of time (i.e., either isochronous or diachronous), a stipulation formalized in the first American Code of Stratigraphic Nomenclature[5] and the International Stratigraphic Guide.[6] This position was reinforced by placement of biostratigraphic units in material categories of the 1983 and 2005 North American Stratigraphic Code.[7] [8] In practice, however, the horizons selected for use as zonal or subzonal boundaries are typically based on the FADs of widely distributed and environmentally tolerant taxa so that the boundaries are not strongly diachronous. It is, after all, commonly time control that is sought in constructing a biostratigraphic framework. Thus, the more closely a zonal boundary approximates an isochron, the better it serves the intended purpose. Consequently, the Cambrian–Ordovician zones and subzones shown in the figures herein are actually biochronozones as defined in the North American Stratigraphic Code.[8] This is not to say that we support the opinion expressed by some (e.g., Ludvigsen et al.;[9] Zalasiewicz et al.[10]) that chronostratigraphic units should be abandoned. Like most stratigraphers (e.g., Ferrusquia-Villafranca et al.[11]), we see the value in the conceptual separation of biostratigraphic, chronostratigraphic, and geochronologic units. The undifferentiated treatment of biozones and biochronozones is merely for concision.
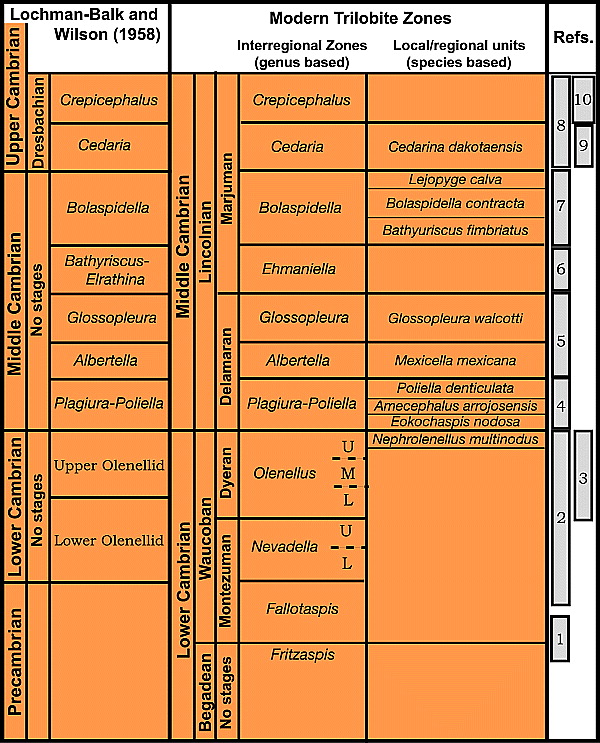
Figure 1 Historical and modern trilobite-based zones within the Lower and the Middle Cambrian platform carbonates of Laurentia and references to recent articles on taxonomic and biostratigraphic refinement for intervals delineated by numbered rectangles in the right column[12]: 1, Hollingsworth;[13] [14] [15] 2, Palmer and Repina;[16] 3, Webster;[17] [18] [19] 4, Sundberg and McCollum;[20] [21] 5, McCollum and Sundberg;[22] 6, Sundberg;[23] [24] 7, Robison;[25] [26] 8, Rasetti;[27] Pratt;[28] 9, Stitt;[29] 10, Stitt and Perfetta.[30]
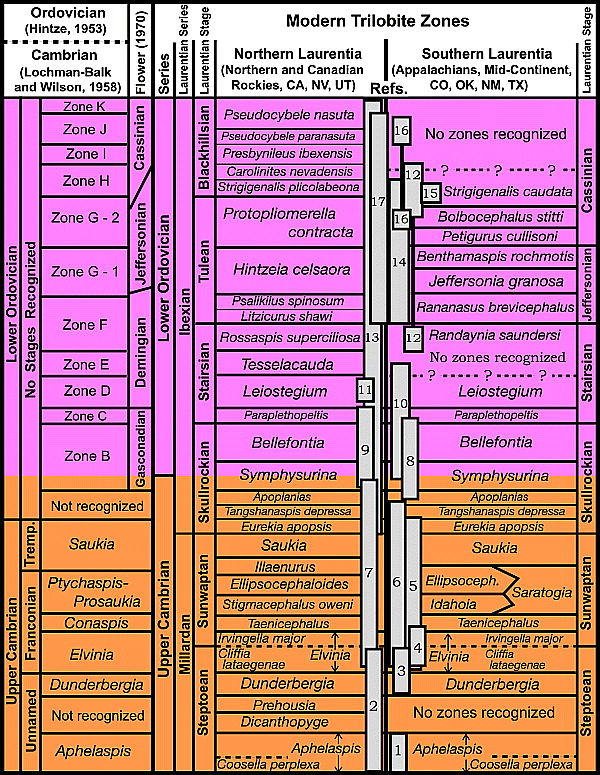
Figure 2 Historical (left column) and modern trilobite-based zones within the Upper Cambrian and the Lower Ordovician platform carbonates of Laurentia with references to articles that describe taxonomic and biostratigraphic refinements for intervals delineated by numbered rectangles near the center of the diagram[12]: 1, Rasetti,[27] Westrop;[31] 2, Palmer,[32] Pratt;[28] 3, Rasetti;[33] 4, Taylor et al.,[34] Loch and Taylor;[35] 5, Longacre;[36] 6, Stitt;[37] 7, Westrop;[38] 8, Stitt;[39] 9, Adrain et al.;[40] 10, Berg and Ross;[41] 11, Pratt;[42] 12, Boyce;[43] 13, Ross et al.,[44] Dean;[45] 14, Loch;[46] 15, Fortey,[47] Boyce,[43] Brett and Westrop;[48] 16, Hu;[49] 17, Adrain et al.[50] Tremp. = Trempealeauan Stage.


Cambrian and Lower Ordovician biochronozones and biomeres[edit]
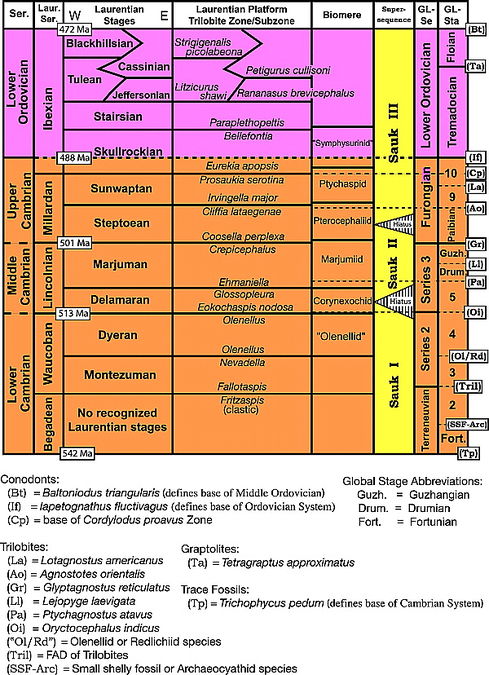
Figure 3 Chronostratigraphic units (series and stages) currently recognized within the Cambrian and the Lower Ordovician deposits (Sauk megasequence) of the great American carbonate bank and related biostratigraphic units (biozones and biomeres). Laurentian series (Laur. Ser.) and stages for the Cambrian are from Palmer;[51] those for the Ordovician are from Ross et al.[44] and Loch.[46] The placement of biomere boundaries follows Taylor.[52] Supersequences are those of Palmer.[53] Global series (GL-Se) and stages (GL-Sta) for the Cambrian are from Shergold and Geyer,[54] Babcock et al.,[55] [56] and Peng et al.;[57] those for the Ordovician are from Webby et al.[58] as modified in Bergstrom et al.[59] Abbreviations for defining taxa/criteria for global series and stage boundaries are provided in small rectangles along right margin and are listed below the figure. Genera undergoing taxonomic revision are shown in quotation marks.[12]
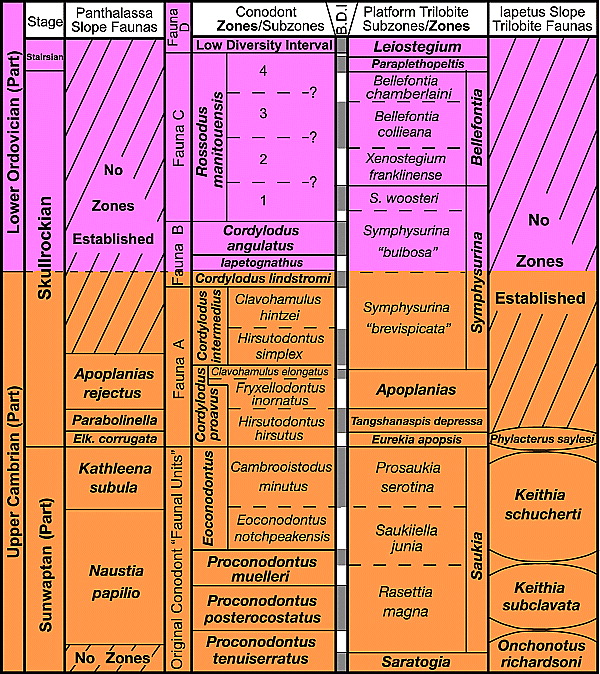
Figure 4 Refined biostratigraphic framework for the uppermost Cambrian and lowest Ordovician of Laurentian North America, showing 25 thin biostratigraphically discriminated intervals (alternating gray and white rectangles in the central column labeled B.D.I.) delineated through a combination of conodont and trilobite zonal or subzonal boundaries through the upper Sunwaptan, Skullrockian, and lowermost Stairsian Stages. Conodont and platform trilobite columns based on Miller et al.[60] with informal numbered subzones in Rossodus manitouensis Zone from Lehnert et al.[61] Zonations for deep-water faunas from southern (Iapetus) and northern (Panthalassa) Laurentian margins from Ludvigsen et al.[62] and Westrop[31] respectively.[12]
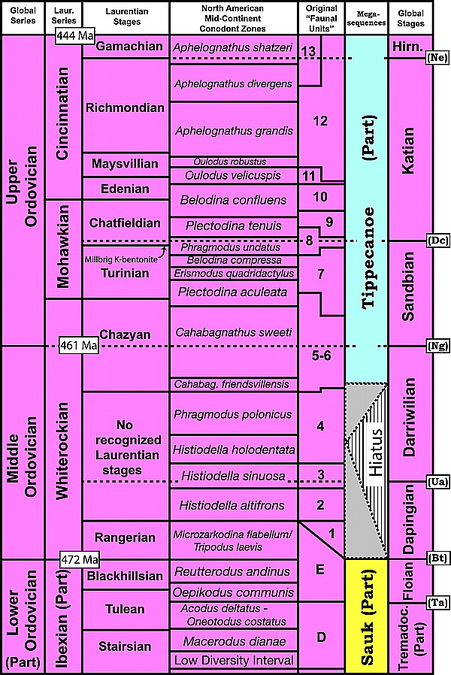
Figure 5 Diagram showing the chronostratigraphic units in the uppermost (Lower Ordovician) part of the Sauk megasequence and the Middle and the Upper Ordovician part of the Tippecanoe megasequence and their relationship to modern North American mid-continent conodont zones, as well as the original alphanumeric conodont faunal units of Ethington and Clark[4] and Sweet et al.[63] Global series and stage names and boundaries are from Webby et al.[58] as modified by Bergstrom et al.[64] Positions and approximate range in ages of the original conodont faunal unit boundaries relative to modern zones are based on a chart compiled by J. E. Repetski and A. G. Harris for use in Poole et al.[65] Abbreviations for species whose first appearance datums (FADs) define the bases of the global stages (shown in rectangles on right margin) include one conodont (Bt = Baltoniodus triangularis) and five graptolites (Ta = Tetragraptus approximatus; Ua = Undulograptus austrodentatus; Ng = Nemagraptus gracilis; Dc = Diplacanthograptus caudatus; and Ne = Normalograptus extraordinarius). Hirn. = Hirnantian Stage; Tremadoc. = Tremadocian Stage.[12] Estimated position of peak of drawdown between Sauk and Tippecanoe megasequences in Histiodella holodentata Zone adopted from Derby et al.[66]



Lochman-Balk and Wilson[1] recognized as their oldest biostratigraphic package a Lower Cambrian Olenellid fauna, which they divided into lower and upper subdivisions based on the appearance of a variety of trilobite genera other than Olenellus. Palmer and Repina[16] delineated three intervals based on the estimated stratigraphic ranges of the Early Cambrian trilobite genera: the Fallotaspis, Nevadella, and Olenellus Zones. This tripartite division formed the basis of the new stages proposed by Palmer[51] for the upper trilobite-bearing part of the Lower Cambrian, which he designated the Waucoban Series (Figure 3). The Fallotaspis Zone and overlying Nevadella Zone together constitute the Montezuman Stage, whereas the upper part comprises the loosely defined Olenellus Zone. Although no well-established fine-scale biozonation is available for the Begadean Series or lower Montezuman Stage (Fallotaspis Zone), recent work by Fritz[67] [68] and Hollingsworth[13] [14] [15] on fallotaspidoid trilobites from western North America has expanded significantly what is known of the oldest trilobites in the Cambrian of Laurentia. The oldest trilobite fauna yet described is that of the Fritzaspis Zone, which was assigned to the uppermost part of the Begadean Series.[15] With few exceptions, the Begadean and lower Montezuman faunas have been recovered from inner-shelf clastic facies. Hence, the biostratigraphic value of the fallotaspidoids for subdivision of carbonate bank deposits is limited at best. In contrast, carbonate deposition spread across wider areas of the Laurentian shelf during the time represented by the overlying Nevadella and Olenellus Zones.
Among the many things revealed by higher resolution biostratigraphic studies through the 1960s and 1970s was that severe extinction events punctuated the evolutionary history of the shallow-marine fauna through the Cambrian. Each event devastated the faunas that inhabited the GACB and the nearshore clastic-dominated environments of the adjacent inner detrital belt. In the aftermath of each extinction, these areas were repopulated by a distinctive group of olenid or olenid-like trilobites (olenimorphs) that migrated inward from deep off-platform environments. This monogeneric replacement fauna invaded all shallow-marine environments across the shelf, reducing the taxonomic and morphologic diversity to a minimum from which it would rise through endemic speciation, augmented by immigration of surviving taxa from distal sites, for the next few million years until the next extinction. The package of strata between the horizons that record the appearances of two successive olenimorph-dominated replacement faunas is a biomere.[32] [69] [70] [71] [72] [73] [52] Five of these stage-level biostratigraphic units have been recognized in the Cambrian System (Figure 3). Each biomere is named for a trilobite family that, although not restricted to that stratigraphic interval, diversified significantly to become a conspicuous component of the faunas that characterize its constituent biozones.
Each biomere-boundary crisis homogenized the platform biota, eliminating the pronounced differences in the taxonomic composition of faunas (biofacies) that simultaneously inhabited different environments before the extinction.[74] [28] [34] [75] The virtual absence of clearly differentiated biofacies in faunas near the biomere boundaries suits them well for use in dividing the Cambrian strata into stages whose boundaries are traceable with exceptional precision across the continent. [76] [28] [51] Conversely, strong biofacies differentiation that developed as faunas recovered from the biomere boundary crises continues to pose a challenge to those attempting to correlate with precision within the body of each biomere. For excellent examples, see the correlation charts provided by Westrop[38] and Hughes and Hesselbo[77] for the upper half of the Sunwaptan Stage (upper part of the Ptychaspid biomere). The two oldest biomere boundaries, at the base and in the middle of the Middle Cambrian, coincide precisely with the bases of the Delamaran and Marjuman Stages (Figure 3). The Middle Cambrian biomere and stage boundaries coincide because the replacement of the diverse preextinction fauna by the minimum diversity olenimorph-dominated fauna appears to have been geologically instantaneous at each of these turnovers. At each of the biomere boundaries in the Upper Cambrian, a thin critical interval[52] with a transitional fauna separates the preextinction fauna from the olenimorph-dominated fauna. Thus, the stage boundaries and biomere boundaries are slightly offset. This contrast in the style of replacement at the Middle versus the Upper Cambrian biomere boundaries and the nature of the transitional fauna in the critical interval are explained more fully below.
The oldest well-documented biomere-like faunal turnover, marked by the extinction of the last olenellid trilobites in the Laurentia at the top of the Olenellus Zone, has been used to define the boundary between the Lower and the Middle Cambrian Series in North America for more than half a century.[1] [51] Unfortunately, this series boundary is unconformable in most areas because of a second-order regression known as the Hawke Bay event.[78] [53] Nonetheless, precise biostratigraphic data from intensive sampling across this boundary in the most complete sections discovered so far confirm that the faunal turnover involved the replacement of a diverse preextinction fauna by one of minimal morphologic and taxonomic diversity. The replacement fauna is overwhelmingly dominated by the generalized ptychopariid trilobite Eokochaspis.[79] [21] The appearance of this minimum-diversity replacement fauna defines both the Olenellid-Corynexochid biomere boundary and the base of the Delamaran Stage and Lincolnian Series.[51] A similar pattern at the base of the overlying Marjuman Stage, where the fauna of the Glossopleura Zone is replaced by a fauna of very low diversity dominated by the generalized ptychpariid Proehmaniella at the base of the Ehmaniella Zone,[23] serves to define the base of the Marjumiid biomere.
As previously noted, a more complex and protracted pattern of faunal turnover has been documented by high-resolution sampling across the three biomere boundaries within the Upper Cambrian: the tops of Marjumiid, Pterocephaliid, and Ptychaspid biomeres (Figure 3). For each of these crises, the diverse preextinction fauna was not replaced immediately by the minimum-diversity replacement fauna. The two are separated stratigraphically by a thin interval with a transitional fauna that is dominated by a surviving opportunistic genus or species from the preextinction fauna, but also includes a few taxa that migrated in from deeper and cooler environments. The base of this critical interval,[52] which Stitt[71] [72] referred to as stage 4 in describing a repeating evolutionary pattern in the Upper Cambrian biomeres, records the extermination of most of the platform trilobites. It can also be recognized by its impact on other faunal groups (e.g., brachiopods and conodonts) and in some cases even in non-Laurentian successions. The breadth of its taxonomic and paleogeographic scope and apparent synchronicity across the entire shelf suit this horizon well for use as a stage boundary within the chronostratigraphic framework. For example, see Miller et al.[80] for a summary of the attributes of the base of the critical interval of the Ptychaspid biomere. Consequently, the bases of the critical intervals that form the uppermost parts of the Marjumiid, Pterocephaliid, and Ptychaspid biomeres are used to define the bases of the Steptoean, Sunwaptan, and Skullrockian Stages, respectively.[76] [51] However, it is the top (not the base) of the critical interval that records the return to minimum diversity with wholesale collapse of platform biofacies and domination of the platform fauna by olenimorphs. For this reason, the top of the critical interval defines the biomere boundary; hence, the Upper Cambrian stage and biomere boundaries are offset stratigraphically from one another by the thickness of the critical interval (Figure 3).
As first noted by Stitt[39] in discussing the Symphysurinid biomere, a similar crisis occurred in the platform trilobite fauna during deposition of the Skullrockian-Stairsian Stage boundary interval in the earliest Ordovician. The pattern of faunal change resembles that documented at the Upper Cambrian biomere or stage boundaries in several respects. There is a thin critical interval (the Paraplethopeltis Zone) dominated by a survivor of the stage-boundary extinction that decimated the diverse fauna of the underlying Bellefontia trilobite Zone (Figure 3 and Figure 4). In addition, a cosmopolitan open-ocean trilobite (Kainella) migrated onto the platform to join the survivors within the critical interval. However, the pattern at the top of the crisis interval (base of the Leiostegium trilobite Zone) differs from the Cambrian biomere boundaries in two critical respects:[81] (1) the trilobite genera that dominate the Paraplethopeltis Zone do not disappear but range upward into the Leiostegium Zone, where they are joined by the species used to define the base of that zone, and (2) consequently, a minimum-diversity olenimorph-dominated replacement fauna comparable to those that typify the Cambrian biomeres never developed. Unlike the Cambrian biomere boundaries, the base of the Leiostegium Zone does not mark the final stage in the extinction process, but records the beginning of the biotic recovery. Because of less severe environmental stress and/or critical zone taxa with higher tolerances, the effect was muted and the virtual depopulation of the platform that occurred during the Cambrian biomere extinction episodes was not repeated.
Regardless of the cause, the change in the dynamics of the extinction-recovery process, from the continent-wide nearly total turnover typical of the Cambrian biomeres to a less complete and more regionalized phenomenon, significantly affected the use of the benthic macrofauna for interregional correlation. Consequently, separate stage nomenclatures for the Lower Ordovician were developed for either side of the Transcontinental Arch.[82] [83] [84] The effects are less pronounced for the lower Ibexian, so the Gasconadian and Demingian Stages of Flower (1964, 1970) have been abandoned in favor of the Skullrockian and Stairsian Stages of Ross et al.[44] which now can be traced from the standard Ibexian Series in Utah into the eastern successions, despite a strong contrast in lithofacies and biofacies between the two regions. However, we retain (Figure 3 and Figure 2) a dual stage terminology for the upper Ibexian, assigning the uppermost Ibexian strata in eastern North America to the Jeffersonian and Cassinian Stages, and restricting the Tulean and Blackhillsian Stages to western North America. However, the bases of the Jeffersonian and Cassinian Stages (rightmost column in Figure 2) differ significantly from those proposed by Flower.[82] The sloping lines used to mark the bases of Flower's Jeffersonian and Cassinian Stages[85] [82] (fourth column from left in Figure 2) are positioned to show the approximate range of diachroneity of these boundaries that resulted from inaccurate correlation between the midcontinent and northern Appalachians.
The conodont zonation for the Upper Cambrian and the Lower Ordovician used herein was developed mostly from extensive work in the eastern Great Basin,[4] [86] [87] [88] [44] but it has been augmented by many studies elsewhere in Laurentian successions (see, for example, Repetski;[89] [90] Ethington and Repetski;[91] Nowlan;[92] Sweet and Bergstrom;[93] Derby et al.;[66] Smith;[94] Ji and Barnes;[95] Harris et al.;[96] Repetski et al.;[97] Landing et al.[98]). The Sauk conodont zonation (Figure 4 and Figure 5) essentially follows that in Ross et al.[44] Two boundaries within this zonation represent profound crises in the conodont faunas of the GACB, both closely associated with major turnovers in the macrofauna: one in the latest Cambrian and the other in the earliest Ordovician. The base of the Cordylodus proavus Zone coincides precisely with the base of the Eurekia apopsis trilobite Zone, which is the base of the critical interval of the Ptychaspid biomere. The extinction of numerous species of trilobites, brachiopods, and conodonts at this horizon, along with the concurrent appearance of many new species of all three groups, makes it one of the most recognizable and traceable boundaries in the lower Paleozoic.[80] Because of its use for correlation throughout Laurentian North America and for recognition of coeval strata deposited on other paleocontinents, Ross et al.[44] selected it to define the base of the Skullrockian Stage and the Ibexian Series.
The other major crisis recorded in the succession of conodont zones within the Sauk megasequence of the Laurentian platform is at the base of the low-diversity interval, just above the base of the Stairsian Stage (Figure 4 and Figure 5). The extinction of many long-ranging conodont species of the underlying Rossodus manitouensis Zone makes this a very significant event in the early evolution of conodonts.[84] [95] Unlike the latest Cambrian crisis at the base of the Skullrockian Stage, in which the conodont and trilobite extinctions apparently were simultaneous, the conodont extinctions at the base of the low-diversity interval postdated the disappearance of the Bellefontia Zone trilobite taxa at the base of the Stairsian Stage.[84] [81] Some conodont species of the R. manitouensis Zone range upward a short distance into the Leiostegium trilobite Zone, producing a very thin overlapping range zone (Figure 4). The presence or absence of these lowermost strata of the Leiostegium Zone that yield conodonts of the R. manitouensis Zone has been used in some studies (e.g., Myrow et al.[99]) to assess the magnitude of the stratigraphic gap in sections where the boundary between the Skullrockian and Stairsian Stages is unconformable.
Middle and Upper Ordovician faunas[edit]
Conodonts have become the dominant tool in zonation and correlation of the Middle and the Upper Ordovician carbonates because of pronounced biofacies- to province-level differentiation that developed in the shelly macrofauna of the GACB during the Middle and the Late Ordovician. The Middle and the Upper Ordovician conodont zones used herein follow those of Webby et al.[58] These zones have evolved from the numbered faunas of Sweet et al.[63] into the succession of named conodont-based chronozones recognized within the composite standard created by Sweet[100] [101] through graphic correlation using species range data from more than 100 Laurentian measured sections. Other important references for conodont faunas of this age include Bergstrom[102] and Harris et al.[103]
Although homotaxial successions of trilobite species useful for local intrabasinal correlation have been documented in some studies (e.g., Fisher;[104]; Ludvigsen;[105] [106] Shaw[107]), no interregional zonation has been developed for these time intervals. Instead, the most recent work on the Middle and the Upper Ordovician shelly macrofaunas has focused mostly on reconstruction of environmental gradients preserved in the lateral arrangement of biofacies or faunas within conodont- or graptolite-based chronozones or between broadly distributed K-bentonite beds or sequence boundaries (e.g., Ludvigsen;[108] Patzkowsky and Holland;[109] [110] Amati and Westrop;[111] Holland and Patzkowsky[112]). The influence of environmental contrasts across the broad carbonate platform created by the Middle and Late Ordovician sea level highstands was compounded further by the development and migration of the Taconic foreland basin in the Appalachians and by associated environmental stresses imposed by episodic tectonics in that region. However, the westward spread of organic-rich deep-water facies did expand the distribution of environments favorable for the preservation of graptolites on the outer margins of the GACB. In the successions that accumulated in such environments, detailed sampling and the integration of biostratigraphic (conodont and graptolite) and geochemical (carbon isotopic and K-bentonite chemistry and age dating) data have produced some of the most finely calibrated chronostratigraphic frameworks and detailed paleoceanographic and tectonostratigraphic models ever constructed for lower Paleozoic strata (e.g., Finney and Bergstrom[113] Mitchell et al.;[114] [115] Ganis and Wise[116]). Such facies are also of particular interest to the petroleum geologist for their importance as source beds.
See also[edit]
References[edit]
- ↑ 1.0 1.1 1.2 Lochman-Balk, C., and J. L. Wilson, 1958, Cambrian biostratigraphy in North America: Journal of Paleontology, v. 32, p. 312–350.
- ↑ Ross Jr., R. J., 1951, Stratigraphy of the Garden City Formation in northeastern Utah, and its trilobite faunas: Peabody Museum of Natural History Bulletin, v. 6, 161 p.
- ↑ Hintze, L. F., 1953, Lower Ordovician trilobites from western Utah: Utah Geological and Mineralogical Survey Bulletin 48, 249 p.
- ↑ 4.0 4.1 4.2 Ethington, R. L., and D. L. Clark, 1971, Lower Ordovician conodonts in North America, in W. C. Sweet and S. M. Bergstrom, eds., Symposium on conodont biostratigraphy: Geological Society of America Memoir 127, p. 63–82.
- ↑ American Commission on Stratigraphic Nomenclature, 1961, Code of stratigraphic nomenclature: AAPG Bulletin, v. 45, p. 645–665.
- ↑ Hedberg, H. D., ed., 1976, International stratigraphic guide: New York, John Wiley and Sons, 200 p.
- ↑ North American Commission on Stratigraphic Nomenclature, 1983, North American stratigraphic code: AAPG Bulletin, v. 67, p. 841–875.
- ↑ 8.0 8.1 North American Commission on Stratigraphic Nomenclature, 2005, North American stratigraphic code: AAPG Bulletin, v. 89, p. 1547–1591, doi:10.1306/07050504129.
- ↑ Ludvigsen, R., S. R. Westrop, B. R. Pratt, P. A. Tuffnell, and G. A. Young, 1986, Dual biostratigraphy: Zones and biofacies: Geoscience Canada, v. 13, p. 139–154.
- ↑ Zalasiewicz, J., et al., 2004, Simplifying the stratigraphy of time: Geology, v. 32, p. 1–4, doi:10.1130/G19920.1.
- ↑ Ferrusquia-Villafranca, I., R. M. Easton, and D. E. Owen, 2009, Do GSSPs render dual time-rock/time classification and nomenclature redundant?: Stratigraphy, v. 6, p. 135–169.
- ↑ 12.0 12.1 12.2 12.3 12.4 Taylor, John F., John E. Repetski, James D. Loch, and Stephen A. Leslie, 2012, Biostratigraphy and chronostratigraphy of the Cambrian–Ordovician great American carbonate bank, in J. R. Derby, R. D. Fritz, S. A. Longacre, W. A. Morgan, and C. A. Sternbach, eds., The great American carbonate bank: The geology and economic resources of the Cambrian–Ordovician Sauk megasequence of Laurentia: AAPG Memoir 98, p. 15–35.
- ↑ 13.0 13.1 Hollingsworth, J. S., 2005, The earliest occurrence of trilobites and brachiopods in the Cambrian of Laurentia: Palaeogeography, Palaeoclimatology, Palaeoecology, v. 220, p. 153–165, doi:10.1016/j.palaeo.2004.08.008.
- ↑ 14.0 14.1 Hollingsworth, J. S., 2006, Holmiidae (Trilobita: Olenellina) of the Montezuman Stage (Early Cambrian) in western Nevada: Journal of Paleontology, v. 80, p. 309–332, doi:10.1666/0022-3360(2006)080[0309:HTOOTM]2.0.CO;2.
- ↑ 15.0 15.1 15.2 Hollingsworth, J. S., 2007, Fallotaspidoid trilobite assemblage (Lower Cambrian) from the Esmeralda Basin (western Nevada, U.S.A.): The oldest trilobites from Laurentia: Memoirs of the Australasian Association of Palaeontologists 32, p. 123–140.
- ↑ 16.0 16.1 Palmer, A. R., and L. N. Repina, 1993, Through a glass darkly: Taxonomy, phylogeny, and biostratigraphy of the Olenellina: Kansas, University of Kansas Paleontological Contributions 3, 35 p.
- ↑ Webster, M., 2003, Olenelloid trilobites of the southern Great Basin, U.S.A., and a refinement of uppermost Dyeran biostratigraphy (abs.): Geological Society of America, Abstracts with Programs, v. 35, p. 166.
- ↑ Webster, M., 2005, Intraspecific variability in Early Cambrian olenelloid trilobites: Implication for biostratigraphy, regional correlation, and phylogeny: Acta Micropalaeontologica Sinica, v. 22, p. 196–197.
- ↑ Webster, M., 2007, Ontogeny and evolution of the Early Cambrian trilobite genus Nephrolenellus (Olenelloidea): Journal of Paleontology, v. 81, p. 1168–1193, doi:10.1666/06-092.1.
- ↑ Sundberg, F. A., and L. B. McCollum, 2003, Trilobites of the Poliella denticulata Biozone (new) of the lower Middle Cambrian of southern Nevada: Journal of Paleontology, v. 77, p. 331–359, doi:10.1666/0022-3360(2003)0772.0.CO;2.
- ↑ 21.0 21.1 Sundberg, F. A., and L. B. McCollum, 2003, Early and mid-Cambrian trilobites from the outer-shelf deposits of Nevada and California, U.S.A.: Palaeontology, v. 46, p. 945–986, doi:10.1111/1475-4983.00328.
- ↑ McCollum, L. B., and F. A. Sundberg, 2007, Cambrian trilobite biozonation of the Laurentian Delamaran Stage in the southern Great Basin, U.S.A.: Implications for global correlations and defining a series 3 global boundary stratotype: Memoirs of the Association of Australasian Palaeontologists 34, p. 147–156.
- ↑ 23.0 23.1 Sundberg, F. A., 1994, Corynexochida and Ptychopariida (Trilobita: Arthropoda) of the Ehmaniella Biozone (Middle Cambrian): Los Angeles County Museum of Natural History, Contributions in Science 446, 137 p.
- ↑ Sundberg, F. A., 2005, The Topazan Stage, a new Laurentian stage (Lincolnian Series–Middle Cambrian): Journal of Paleontology, v. 79, p. 63–71, doi:10.1666/0022-3360(2005)0792.0.CO;2.
- ↑ Robison, R. A., 1964, Late Middle Cambrian faunas from western Utah: Journal of Paleontology, v. 38, p. 510–566.
- ↑ Robison, R. A., 1976, Middle Cambrian trilobite biostratigraphy of the Great Basin, in R. A. Robison and A. J. Rowell, eds., Paleontology and depositional environments: Cambrian of western North America: Brigham Young University Geology Studies, v. 23, p. 39–50.
- ↑ 27.0 27.1 Rasetti, F., 1965, Upper Cambrian trilobite faunas of northeastern Tennessee: Smithsonian Miscellaneous Collections, v. 148, 127 p.
- ↑ 28.0 28.1 28.2 28.3 Pratt, B. R., 1992, Trilobites of the Marjuman and Steptoean Stages (Upper Cambrian), Rabbitkettle Formation, southern Mackenzie Mountains, northwest Canada: Palaeontographica Canadiana, v. 9, 179 p.
- ↑ Stitt, J. H., 1998, Trilobites from the Cedarina dakotaensis Zone, lowermost part of the Deadwood Formation (Marjuman Stage, Upper Cambrian), Black Hills, South Dakota: Journal of Paleontology, v. 2, p. 1030–1046.
- ↑ Stitt, J. H., and P. J. Perfetta, 2000, Trilobites, biostratigraphy, and lithostratigraphy of the Crepicephalus and Aphelaspis Zones, Lower Deadwood Formation (Marjuman and Steptoean Stages, Upper Cambrian), Black Hills, South Dakota: Journal of Paleontology, v. 74, p. 199–223, doi:10.1666/0022-3360(2000)0742.0.CO;2.
- ↑ 31.0 31.1 Westrop, S. R., 1992, Upper Cambrian (Marjuman-Steptoean) trilobites from the Port Au Port Group, western Newfoundland: Journal of Paleontology, v. 66, p. 228–255.
- ↑ 32.0 32.1 Palmer, A. R., 1965, Trilobites of the Late Cambrian Pterocephaliid biomere in the Great Basin, United States: U.S. Geological Survey Professional Paper 493, 105 p.
- ↑ Rasetti, F., 1961, Dresbachian and Franconian trilobites of the Conococheague and Frederick limestones of the central Appalachians: Journal of Paleontology, v. 35, p. 104–124.
- ↑ 34.0 34.1 Taylor, J. F., J. D. Loch, and P. J. Perfetta, 1999, Trilobite faunas from Upper Cambrian reefs in the central Appalachians: Journal of Paleontology, v. 73, p. 326–336.
- ↑ Loch, J. D., and J. F. Taylor, 2004, New trilobite taxa from Upper Cambrian microbial reefs in the central Appalachians: Journal of Paleontology, v. 78, p. 591–602, doi:10.1666/0022-3360(2004)0782.0.CO;2.
- ↑ Longacre, S. A., 1970, Trilobites of the Upper Cambrian Ptychaspid biomere, Wilberns Formation, central Texas: Paleontological Society Memoir 4, Journal of Paleontology, v. 44 supp., 70 p.
- ↑ Stitt, J. H., 1971b, Late Cambrian and earliest Ordovician trilobites: Timbered Hills and Lower Arbuckle Groups, western Arbuckle Mountains, Murray County, Oklahoma: Oklahoma Geological Survey Bulletin 110, 83 p.
- ↑ 38.0 38.1 Westrop, S. R., 1986, Trilobites of the Upper Cambrian Sunwaptan Stage, southern Canadian Rocky Mountains, Alberta: Palaeontographica Canadiana, v. 3, 179 p.
- ↑ 39.0 39.1 Stitt, J. H., 1983, Trilobite biostratigraphy and lithostratigraphy of the McKenzie Hill Limestone (Lower Ordovician), Wichita and Arbuckle Mountains, Oklahoma: Oklahoma Geological Survey Bulletin, v. 134, 54 p.
- ↑ Adrain, J. M., D.-C. Lee, S. R. Westrop, B. D. E. Chatterton, and E. Landing, 2003, Classification of the trilobite subfamilies Hystricurinae and Hintzecurinae subfam. nov., with new genera from the Lower Ordovician (Ibexian) of Idaho and Utah: Memoirs of the Queensland Museum 48, p. 543–586.
- ↑ Berg, R. R., and R. J. Ross Jr., 1959, Trilobites from the Peerless and Manitou Formations, Colorado: Journal of Paleontology, v. 33, p. 106–119.
- ↑ Pratt, B. R., 1988, An Ibexian (Early Ordovician) trilobite faunule from the type section of the Rabbitkettle Formation (southern Mackenzie Mountains, Northwest Territories): Canadian Journal of Earth Sciences, v. 25, p. 1595–1607, doi:10.1139/e88-152.
- ↑ 43.0 43.1 Boyce, W. D., 1989, Early Ordovician trilobite faunas of the Boat Harbour and Catoche Formations (St. George Group), western Newfoundland: Newfoundland Department of Mines and Energy, Geological Survey Branch, Report 89-22 m, 169 p.
- ↑ 44.0 44.1 44.2 44.3 44.4 44.5 Ross Jr., R. J., L. F. Hintze, R. L. Ethington, J. F. Miller, M. E. Taylor, J. E. Repetski, J. Sprinkle, and T. E. Guensburg, 1997, The Ibexian, lowermost series in the North American Ordovician, in M. E. Taylor, ed., Early Paleozoic biochronology of the Great Basin, western United States: U.S. Geological Survey Professional Paper 1579-A, p. 1–50.
- ↑ Dean, W. T., 1989, Trilobites from the Survey Peak, Outram, and Skoki Formations (Upper Cambrian–Lower Ordovician) at Wilcox Pass, Jasper National Park, Alberta: Geological Survey of Canada Bulletin 389, 141 p.
- ↑ 46.0 46.1 Loch, J. D., 2007, Trilobite biostratigraphy and correlation of the Lower Ordovician Kindblade Formation of Carter and Kiowa counties, Oklahoma: Oklahoma Geological Survey Bulletin 149, 157 p.
- ↑ Fortey, R. A., 1979, Early Ordovician trilobites from the Catoche Formation (St. George Group), western Newfoundland: Geological Survey of Canada Bulletin, v. 321, p. 61–114.
- ↑ Brett, K. D., and S. R. Westrop, 1996, Trilobites of the Lower Ordovician (Ibexian) Fort Cassin Formation, Champlain Valley region, New York State and Vermont: Journal of Paleontology, v. 70, p. 408–427.
- ↑ Hu, C.-H., 1963, Some Lower Ordovician trilobites from Franklin Mountains, Texas: Transactions Proceedings of the Paleontological Society of Japan, no. 51, p. 86–90.
- ↑ Adrain, J. M., N. E. B. McAdams, and S. R. Westrop, 2009, Trilobite biostratigraphy and revised bases of the Tulean and Blackhillsian Stages of the Ibexian Series, Lower Ordovician, western United States, in J. R. Laurie, ed., Cambro–Ordovician studies IV: Memoirs of the Association of Australasian Palaeontologists 34, p. 541–610.
- ↑ 51.0 51.1 51.2 51.3 51.4 51.5 Palmer, A. R., 1998, A proposed nomenclature for stages and series for the Cambrian of Laurentia: Canadian Journal of Earth Sciences, v. 35, p. 323–328.
- ↑ 52.0 52.1 52.2 52.3 Taylor, J. F., 2006, History and status of the biomere concept, in J. R. Paterson and J. R. Laurie, eds., Cambro–Ordovician studies II: Memoirs of the Association of Australasian Palaeontologists 32, p. 247–265.
- ↑ 53.0 53.1 Palmer, A. R., 1981, Subdivision of the Sauk sequence, in M. E. Taylor, ed., Second International Symposium on the Cambrian System: U.S. Geological Survey Open-File Report 81-743, p. 160–162.
- ↑ Shergold, J. H., and G. Geyer, 2003, The subcommisssion on Cambrian stratigraphy: The status quo: Geologica Acta, v. 1, p. 5–9.
- ↑ Babcock, L. E., S. Peng, G. Geyer, and J. H. Shergold, 2005, Changing perspectives on Cambrian chronostratigraphy and progress toward subdivision of the Cambrian System: Geosciences Journal, v. 9, p. 101–106, doi:10.1007/BF02910572.
- ↑ Babcock, L. E., R. A. Robison, M. N. Rees, S. Peng, and M. R. Saltzman, 2007, The global boundary stratotype section and point (GSSP) of the Drumian Stage (Cambrian) in the Drum Mountains, Utah, U.S.A.: Episodes, v. 30, p. 84–94.
- ↑ Peng, S., L. E. Babcock, R. A. Robison, L. Huanling, M. N. Rees, and M. R. Saltzman, 2004, Global Standard Stratotype Section and Point (GSSP) of the Furongian Series and Paibian Stage (Cambrian): Lethaia, v. 37, p. 365–379, doi:10.1080/00241160410002081.
- ↑ 58.0 58.1 58.2 Webby, B. D., R. A. Cooper, S. M. Bergstrom, and F. Paris, 2004, Stratigraphic framework and time slices, in B. D. Webby, F. Paris, M. L Droser, and I. G. Percival, eds., The great Ordovician biodiversification event: New York, Columbia University Press, p. 41–47.
- ↑ Bergstrom, S. M., S. C. Finney, X. Chen, D. Goldman, and S. A. Leslie, 2006, Three new Ordovician global stage names: Lethaia, v. 39, p. 287–288, doi:10.1080/00241160600847439.
- ↑ Miller, J. F., K. R. Evans, J. D. Loch, R. L. Ethington, J. H. Stitt, L. Holmer, and L. E. Popov, 2003, Stratigraphy of the Sauk III interval (Cambrian–Ordovician) in the Ibex area, western Millard County, Utah and central Texas: Brigham Young University Geology Studies, v. 47, p. 23–118.
- ↑ Lehnert, O., J. F. Miller, S. A. Leslie, J. E. Repetski, and R. L. Ethington, 2005, Cambro–Ordovician sea level fluctuations and sequence boundaries: The missing record and the evolution of new taxa: The Palaeontological Association, Special Papers in Palaeontology 73, p. 117–134.
- ↑ Ludvigsen, R., S. R. Westrop, and C. H. Kindle, 1989, Sunwaptan (Upper Cambrian) trilobites from the Cow Head Group, western Newfoundland, Canada: Palaeontographica Canadiana 6, 175 p.
- ↑ 63.0 63.1 Sweet, W. C., R. L. Ethington, and C. R. Barnes, 1971, North American Middle and Upper Ordovician conodont faunas, in W. C. Sweet and S. M. Bergstrom, eds., Symposium on conodont biostratigraphy: Geological Society of America Memoir 127, p. 163–193.
- ↑ Bergstrom, S. M., X. Chen, J. C. Gutierrez-Marco, and A. Dronov, 2008, The new chronostratigraphic classification of the Ordovician System and its relations to major regional series and stages and to δ13C chemostratigraphy: Lethaia, v. 41, p. 1–11, doi:10.1111/j.1502-3931.2007.00039.x.
- ↑ Poole, F. G., J. H. Stewart, J. E. Repetski, A. G. Harris, R. J. Ross Jr., K. B. Ketner, R. Amaya-Martinez, and J. M. Morales-Ramirez, 1995, Ordovician carbonate-shelf rocks of Sonora, Mexico, in J. D. Cooper, M. L. Droser, and S. C. Finney, eds., Ordovician odyssey: Short papers of the seventh international symposium on the Ordovician System: Pacific Section SEPM 77, p. 267–275.
- ↑ 66.0 66.1 Derby, J. R., et al., 1991, Biostratigraphy of the Timbered Hills, Arbuckle, and Simpson groups, Cambrian and Ordovician, Oklahoma: A review of correlation tools and techniques available to the explorationist, in K. Johnson, ed., Late Cambrian–Ordovician geology of the southern mid-continent: Oklahoma Geological Survey Circular 92, p. 15–41.
- ↑ Fritz, W. H., 1972, Lower Cambrian trilobites from the Sekwi Formation type section, Mackenzie Mountains, northwestern Canada: Geological Survey of Canada Bulletin 212, 90 p.
- ↑ Fritz, W. H., 1993, New Lower Cambrian olenelloid trilobite genera Cirquella and Geraldinella from southwestern Canada: Journal of Paleontology, v. 67, p. 856–868.
- ↑ Palmer, A. R., 1965b, Biomere: A new kind of biostratigraphic unit: Journal of Paleontology, v. 39, p. 149–153.
- ↑ Palmer, A. R., 1984, The biomere problem: Evolution of an idea: Journal of Paleontology, v. 58, p. 599–611.
- ↑ 71.0 71.1 Stitt, J. H., 1971a, Repeating evolutionary pattern in Late Cambrian biomeres: Journal of Paleontology, v. 45, p. 178–181.
- ↑ 72.0 72.1 Stitt, J. H., 1975, Adaptive radiation, trilobite paleoecology, and extinction, Ptychaspid biomere, Late Cambrian of Oklahoma: Fossils and Strata, v. 4, p. 381–390.
- ↑ Stitt, J. H., 1977, Late Cambrian and earliest Ordovician trilobites, Wichita Mountains area, Oklahoma: Oklahoma Geological Survey Bulletin, v. 124, 79 p.
- ↑ Ludvigsen, R., and S. R. Westrop, 1983, Trilobite biofacies of the Cambrian-Ordovician boundary interval in northern North America: Alcheringa, v. 7, p. 301–319, doi:10.1080/03115518308619614.
- ↑ Westrop, S. R., and M. B. Cuggy, 1999, Comparative paleoecology of Cambrian trilobite extinctions: Journal of Paleontology, v. 73, p. 337–354.
- ↑ 76.0 76.1 Ludvigsen, R., and S. R. Westrop, 1985, Three new Upper Cambrian stages for North America: Geology, v. 13, p. 139–143, doi:10.1130/0091-7613(1985)132.0.CO;2.
- ↑ Hughes, N. C., and S. P. Hesselbo, 1997, Stratigraphy and sedimentology of the St. Lawrence Formation, Upper Cambrian of the northern Mississippi Valley: Milwaukee Public Museum Contributions in Biology and Geology 91, 50 p.
- ↑ Palmer, A. R., and N. P. James, 1979, The Hawke Bay event: A circum-Iapetus regression near the Lower Middle Cambrian boundary, in D. R. Wones, ed., The Caledonides in the U.S.A.: Proceedings, International Geological Correlation Programme, project 27: Caledonide orogen: Virginia Polytechnic Institute and State University, Department of Geological Sciences Memoir 2, p. 15–18.
- ↑ Sundberg, F. A., and L. B. McCollum, 2000, Ptychopariid trilobites of the Lower–Middle Cambrian boundary interval, Pioche Shale, southeastern Nevada: Journal of Paleontology, v. 74, p. 604–630, doi:10.1666/0022-3360(2000)074lt0604:PTOTLMgt2.0.CO;2.
- ↑ 80.0 80.1 Miller, J. F., R. L. Ethington, K. R. Evans, L. E. Holmer, J. D. Loch, L. E. Popov, J. E. Repetski, R. L. Ripperdan, and J. F. Taylor, 2006, Proposed stratotype for the base of the highest Cambrian stage at the first appearance datum of Cordylodus andresi, Lawson Cove section, Utah, U.S.A.: Paleoworld, v. 15, p. 384–405, doi:10.1016/j.palwor.2006.10.017.
- ↑ 81.0 81.1 Taylor, J. F., D. K. Brezinski, J. E. Repetski, and N. M. Welsh, 2009a, The Adamstown submergence event: Faunal and sedimentological record of a Late Cambrian transgression in the Appalachian Region, in J. R. Laurie, ed., Cambro–Ordovician studies IV: Memoirs of the Association of Australasian Palaeontologists 34, p. 147–156.
- ↑ 82.0 82.1 82.2 Flower, R. H., 1970, Early Paleozoic of New Mexico and the El Paso region, revision 2: New Mexico Bureau of Mines and Mineral Resources, reprint series, 44 p.
- ↑ Ross Jr., R. J., 1976, Ordovician sedimentation in the western United States, in M. G. Bassett, ed., The Ordovician System: Proceedings of a Palaeontological Association symposium: Cardiff, University of Wales Press and National Museum of Wales, p. 73–105.
- ↑ 84.0 84.1 84.2 Ethington, R. L., K. M. Engel, and K. L. Elliott, 1987, An abrupt change in conodont faunas in the Lower Ordovician of the mid-continent province, in R. J. Aldridge, ed., Paleobiology of conodonts: Chichester, United Kingdom, Ellis Horwood Limited, p. 111–127.
- ↑ Flower, R. H., 1964, The nautiloid order Ellesmeroceratida (Cephalopoda): New Mexico Bureau of Mines and Mineral Resources Memoir 12, 234 p.
- ↑ Ethington, R. L., and D. L. Clark, 1982, Lower and Middle Ordovician conodonts from the Ibex area, western Millard County, Utah: Brigham Young University Geology Studies, v. 28, 155 p. (imprint 1981).
- ↑ Miller, J. F., 1980, Taxonomic revisions of some Upper Cambrian and Lower Ordovician conodonts with comments on their evolution: University of Kansas, Paleontological Contributions 99, 43 p.
- ↑ Miller, J. F., 1988, Conodonts as biostratigraphic tools for redefinition and correlation of the Cambrian-Ordovician boundary: Geological Magazine, v. 125, p. 349–362, doi:10.1017/S0016756800013029.
- ↑ Repetski, J. E., 1977, Early Ordovician (Canadian) conodonts from New York (abs.): Geological Society of America Abstracts with Programs, v. 9, no. 5, 647 p.
- ↑ Repetski, J. E., 1985, Conodont biostratigraphy of the Knox Group at the Thorn Hill and River Ridge sections, northeastern Tennessee, in K. R. Walker, ed., The geologic history of the Thorn Hill Paleozoic section (Cambrian–Mississippian), eastern Tennessee: University of Tennessee Department of Geological Sciences Studies in Geology, no. 10, p. 25–31.
- ↑ Ethington, R. L., and J. E. Repetski, 1984, Paleobiogeographic distribution of Early Ordovician conodonts in central and western United States, in D. L. Clark, ed., Conodont biofacies and provincialism: Geological Society of America, Special Paper 196, p. 89–101.
- ↑ Nowlan, G. S., 1985, Cambrian–Ordovician conodonts from the Franklinian miogeosyncline, Canadian Arctic Islands: Journal of Paleontology, v. 59, p. 96–122.
- ↑ Sweet, W. C., and S. M. Bergstrom, 1986, Conodonts and biostratigraphic correlation: Annual Reviews of Earth and Planetary Science, v. 14, p. 85–112.
- ↑ Smith, M. P., 1991, Early Ordovician conodonts of east and north Greenland: Meddelelser om Gronland 26, 81 p.
- ↑ 95.0 95.1 Ji, Z., and C. R. Barnes, 1994, Lower Ordovician conodonts of the St. George Group, Port au Port Peninsula, western Newfoundland, Canada: Palaeontographica Canadiana, v. 11, 149 p.
- ↑ Harris, A. G., J. A. Dumoulin, J. E. Repetski, and C. Carter, 1995, Correlation of Ordovician rocks of northern Alaska: in J. D. Cooper, M. L. Droser, and S. C. Finney, eds., Ordovician odyssey: Short papers for the 7th International symposium on the Ordovician System: Pacific Section SEPM Publication 77, p. 21–26.
- ↑ Repetski, J. E., A. G. Harris, and N. R. Stamm, 1995, An overview of conodonts from New Jersey: in J. E. B. Baker, ed., Contributions to the paleontology of New Jersey: The Geological Association of New Jersey, v. XII, p. 191–208.
- ↑ Landing, E., S. R. Westrop, and L. Van Aller Hernick, 2003, Uppermost Cambrian–Lower Ordovician faunas and Laurentian platform sequence stratigraphy, eastern New York and Vermont: Journal of Paleontology, v. 77, p. 78–98, doi:10.1666/0022-3360(2003)0772.0.CO;2.
- ↑ Myrow, P. M., J. F. Taylor, J. F. Miller, R. L. Ethington, R. L. Ripperdan, and J. Allen, 2003, Fallen arches: Dispelling myths concerning Cambrian and Ordovician paleogeography of the Rocky Mountain region: Geological Society of America Bulletin, v. 115, p. 695–713, doi:10.1130/0016-7606(2003)1152.0.CO;2.
- ↑ Sweet, W. C., 1984, Graphic correlation of upper Middle and Upper Ordovician rocks, North American mid-continent province, U.S.A., in D. L. Bruton, ed., Aspects of the Ordovician System: Paleontological contributions from the University of Oslo, no. 295, p. 23–35.
- ↑ Sweet, W. C., 1995, Graphic assembly of a conodont-based composite standard for the Ordovician System of North America, in K. O. Mann and H. R. Lane, eds., Graphic correlation: SEPM Special Publication 53, p. 139–150.
- ↑ Bergstrom, S. M., 1971, Conodont biostratigraphy of the Middle and Upper Ordovician of Europe and eastern North America, in W. C. Sweet and S. M. Bergstrom, eds., Symposium on conodont biostratigraphy: Geological Society of America Memoir 127, p. 83–164.
- ↑ Harris, A. G., S. M. Bergstrom, R. L. Ethington, and R. J. Ross Jr., 1979, Aspects of Middle and Upper Ordovician conodont biostratigraphy of carbonate facies in Nevada and southeast California and comparison with some Appalachian successions, in C. A. Sandberg and D. L. Clark, eds., Conodont biostratigraphy of the Great Basin and Rocky Mountains: Brigham Young University Geology Studies, v. 26, p. 7–43.
- ↑ Fisher, D. W., 1977, Correlation of the Hadrynian, Cambrian, and Ordovician rocks in New York State: New York State Museum, Map and Chart Series 25, 75 p.
- ↑ Ludvigsen, R., 1977, The Ordovician trilobite Ceraurinus Barton in North America: Journal of Paleontology, v. 51, p. 559–572.
- ↑ Ludvigsen, R., 1979, A trilobite zonation of Middle Ordovician rocks, southwestern district of Mackenzie: Geological Survey of Canada Bulletin 312, 99 p.
- ↑ Shaw, F. C., 1991, Viola Group (Ordovician, Oklahoma) cryptolithinid trilobites: Biostratigraphy and taxonomy: Journal of Paleontology, v. 65, p. 919–935.
- ↑ Ludvigsen, R., 1978, Middle Ordovician trilobite biofacies, southern Mackenzie Mountains, in C. R. Stelck and B. D. E. Chatterton, eds., Western and arctic Canadian biostratigraphy: Geological Association of Canada Special Paper 18, p. 1–37.
- ↑ Patzkowsky, M. E., and S. M. Holland, 1996, Extinction, invasion, and sequence stratigraphy: Patterns of faunal change in the Middle and Upper Ordovician of the eastern United States: Geological Society of America Special Paper 306, p. 131–142.
- ↑ Patzkowsky, M. E., and S. M. Holland, 1999, Biofacies replacement in a sequence-stratigraphic framework: Middle and Upper Ordovician of the Nashville Dome, Tennessee, U.S.A.: PALAIOS, v. 14, p. 301–317, doi:10.2307/3515459.
- ↑ Amati, L. M., and S. R. Westrop, 2006, Sedimentary facies and trilobite biofacies along an Ordovician shelf to basin gradient, Viola Group, south-central Oklahoma: PALAIOS, v. 21, p. 516–529, doi:10.2110/palo.2006.p06-069.
- ↑ Holland, S. M., and M. E. Patzkowsky, 2007, Gradient ecology of a biotic invasion: Biofacies of the type Cincinnatian Series (Upper Ordovician), U.S.A.: PALAIOS, v. 22, p. 392–407, doi:10.2110/palo.2006.p06-066r.
- ↑ Finney, S. C., and S. M. Bergstrom, 1986, Biostratigraphy of the Ordovician Nemagraptus gracilis Zone, in C. P. Hughes and R. B. Rickards, eds., Palaeoecology and biostratigraphy of graptolites: Geological Society Special Publication 20, p. 47–59.
- ↑ Mitchell, C. E., D. Goldman, J. W. Delano, S. D. Samson, and S. M. Bergstrom, 1994, Temporal and spatial distribution of biozones and facies relative to geochemically correlated K-bentonites in the Middle Ordovician Taconic foredeep: Geology, v. 22, p. 715–718, doi:10.1130/0091-7613(1994)0222.3.CO;2.
- ↑ Mitchell, C. E., S. Adhya, S. M. Bergstrom, M. P. Joy, and J. W. Delano, 2004, Discovery of the Ordovician Millbrig K-bentonite Bed in the Trenton Group of New York State: Implications for regional correlation and sequence stratigraphy in eastern North America: Palaeogeography, Palaeoclimatology, Palaeoecology, v. 210, p. 331–346, doi:10.1016/j.palaeo.2004.02.037.
- ↑ Ganis, R. G., and D. U. Wise, 2008, Taconic events in Pennsylvania: Datable phases of a 20 m.y. orogeny: American Journal of Science, v. 308, p. 16–183.
External links[edit]
| find literature about Great American carbonate bank, biostratigraphy |